
Aileron de poisson -Fish fin
.png)
(1) nageoires pectorales (appariées), (2) nageoires pelviennes (appariées), (3) nageoire dorsale ,
(4) nageoire adipeuse, (5) nageoire anale, (6) caudale ( nageoire caudale
Les nageoires sont des caractéristiques anatomiques distinctives composées d' épines osseuses ou de rayons dépassant du corps d'un poisson . Ils sont recouverts de peau et réunis soit de manière palmée, comme on le voit chez la plupart des poissons osseux , soit comme une nageoire , comme on le voit chez les requins . En dehors de la queue ou de la nageoire caudale , les nageoires des poissons n'ont aucun lien direct avec la colonne vertébrale et ne sont soutenues que par des muscles . Leur fonction principale est d'aider les poissons à nager .
Les nageoires situées à différents endroits sur le poisson servent à différentes fins telles qu'avancer, tourner, garder une position verticale ou s'arrêter. La plupart des poissons utilisent des nageoires pour nager, les poissons volants utilisent des nageoires pectorales pour glisser et les poissons grenouilles les utilisent pour ramper. Les palmes peuvent également être utilisées à d'autres fins; les requins mâles et les moustiques utilisent une nageoire modifiée pour délivrer du sperme, les requins -renards utilisent leur nageoire caudale pour étourdir leurs proies, les poissons-pierres de récif ont des épines dans leurs nageoires dorsales qui injectent du venin, les baudroies utilisent la première épine de leur nageoire dorsale comme une canne à pêche pour attirer leurs proies , et les balistes évitent les prédateurs en se faufilant dans les crevasses coralliennes et en utilisant des épines dans leurs nageoires pour se verrouiller en place.
Types de palmes
Pour chaque type de nageoire, il existe un certain nombre d'espèces de poissons chez lesquelles cette nageoire particulière a été perdue au cours de l'évolution.
| Nageoires pectorales |

|
Les nageoires pectorales appariées sont situées de chaque côté, généralement maintenues repliées juste derrière l'opercule, et sont homologues aux membres antérieurs des tétrapodes .
|
|---|---|---|
|
Nageoires pelviennes (nageoires ventrales) |

|
Les nageoires pelviennes ou ventrales appariées sont généralement situées ventralement en dessous et derrière les nageoires pectorales, bien que dans de nombreuses familles de poissons, elles puissent être positionnées devant les nageoires pectorales (par exemple les cabillauds). Ils sont homologues aux membres postérieurs des tétrapodes . La nageoire pelvienne aide le poisson à monter ou descendre dans l'eau, à tourner brusquement et à s'arrêter rapidement.
|
| Nageoire dorsale |
 |

Nageoire dorsale d'un chevesne ( Leuciscus cephalus )
Les nageoires dorsales sont situées sur le dos. Un poisson peut avoir jusqu'à trois nageoires dorsales. Les nageoires dorsales servent à protéger le poisson contre le roulis et l'aident dans les virages et les arrêts brusques.
|
| Nageoire anale/cloacale |

|
La nageoire anale/cloacale est située sur la surface ventrale derrière l' anus / cloaque . Cette palme sert à stabiliser le poisson pendant la nage. |
| Nageoire adipeuse |
 |
La nageoire adipeuse est une nageoire molle et charnue située à l'arrière de la nageoire dorsale et juste en avant de la nageoire caudale. Il est absent de nombreuses familles de poissons, mais présent dans neuf des 31 ordres eutéléostéens ( Percopsiformes , Myctophiformes , Aulopiformes , Stomiiformes , Salmoniformes , Osmeriformes , Characiformes , Siluriformes et Argentiniformes ). Les représentants célèbres de ces ordres sont le saumon , les characidés et le poisson- chat .
La fonction de la nageoire adipeuse est un mystère. Il est fréquemment coupé pour marquer les poissons élevés en écloserie, bien que les données de 2005 aient montré que les truites dont la nageoire adipeuse a été retirée ont une fréquence de battement de queue 8% plus élevée. Des informations supplémentaires publiées en 2011 suggèrent que la nageoire peut être vitale pour la détection et la réponse à des stimuli tels que le toucher, le son et les changements de pression. Des chercheurs canadiens ont identifié un réseau de neurones dans la nageoire, indiquant qu'il a probablement une fonction sensorielle, mais ne savent toujours pas exactement quelles sont les conséquences de son retrait. Une étude comparative de 2013 indique que la nageoire adipeuse peut se développer de deux manières différentes. L'une est la voie de type salmoniforme, où la nageoire adipeuse se développe à partir du pli larvaire en même temps et de la même manière directe que les autres nageoires médianes. L'autre est la voie de type characiforme, où la nageoire adipeuse se développe tardivement après que le pli de la nageoire larvaire a diminué et que les autres nageoires médianes se sont développées. Ils affirment que l'existence du type de développement characiforme suggère que la nageoire adipeuse n'est pas "juste un reste de pli de nageoire larvaire" et est incompatible avec l'idée que la nageoire adipeuse manque de fonction. Des recherches publiées en 2014 indiquent que la nageoire adipeuse a évolué à plusieurs reprises dans des lignées distinctes . |
| Nageoire caudale (nageoire caudale) |
 
|
La nageoire caudale est la nageoire caudale (du latin cauda signifiant queue), située à l'extrémité du pédoncule caudal et sert à la propulsion. Voir locomotion corps-nageoire caudale .
(A) - Hétérocerque signifie que les vertèbres s'étendent dans le lobe supérieur de la queue, ce qui la rend plus longue (comme chez les requins ). C'est le contraire d'hypocerque.
(B) - Protocercal signifie que les vertèbres s'étendent jusqu'au bout de la queue et que la queue est symétrique mais non élargie (comme chez les premiers poissons et les cyclostomes , et un précurseur plus primitif chez les lancelettes ) (C) - Homocercal où la nageoire apparaît superficiellement symétrique mais en fait les vertèbres s'étendent sur une très courte distance dans le lobe supérieur de la nageoire (D) - Diphycerque signifie que les vertèbres s'étendent jusqu'au bout de la queue et que la queue est symétrique et élargie (comme chez le bichir , le dipneuste , la lamproie et le cœlacanthe ). La plupart des poissons paléozoïques avaient une queue hétérocerque diphycerque. La plupart des poissons modernes ( téléostéens ) ont une queue homocerque. Ceux-ci apparaissent dans une variété de formes et peuvent apparaître :
|
| Quille caudale Finlets |
_diagram_cropped.GIF)
|
Certains types de poissons à nage rapide ont une quille caudale horizontale juste en avant de la nageoire caudale. Tout comme la quille d'un navire, il s'agit d'une crête latérale sur le pédoncule caudal, généralement composée d'écailles (voir ci-dessous), qui assure la stabilité et le soutien de la nageoire caudale. Il peut y avoir une seule paire de quilles, une de chaque côté, ou deux paires au-dessus et au-dessous.
Les pinnules sont de petites nageoires, généralement derrière les nageoires dorsale et anale (chez les bichirs , il n'y a que des pinnules sur la face dorsale et pas de nageoire dorsale). Chez certains poissons comme le thon ou le balaou , ils sont sans rayons, non rétractables et se trouvent entre la dernière nageoire dorsale et/ou anale et la nageoire caudale. |
Poissons osseux

Les poissons osseux forment un groupe taxonomique appelé Osteichthyes. Ils ont des squelettes en os et peuvent être comparés aux poissons cartilagineux qui ont des squelettes en cartilage . Les poissons osseux sont divisés en poissons à nageoires rayonnées et à nageoires lobes . La plupart des poissons sont à nageoires rayonnées, un groupe extrêmement diversifié et abondant composé de plus de 30 000 espèces. C'est la plus grande classe de vertébrés qui existe aujourd'hui. Dans un passé lointain, les poissons à nageoires lobes étaient abondants. De nos jours, ils sont principalement éteints, avec seulement huit espèces vivantes. Les poissons osseux ont des épines et des rayons de nageoire appelés lépidotriches. Ils ont généralement des vessies natatoires , ce qui permet au poisson de créer un équilibre neutre entre couler et flotter sans avoir à utiliser ses nageoires. Cependant, les vessies natatoires sont absentes chez de nombreux poissons, notamment chez les poumons , qui sont les seuls poissons à avoir conservé le poumon primitif présent chez l'ancêtre commun des poissons osseux à partir desquels les vessies natatoires ont évolué. Les poissons osseux ont également un opercule , qui les aide à respirer sans avoir à utiliser des nageoires pour nager.
Lobe-nageoires

Les poissons à nageoires lobes forment une classe de poissons osseux appelés Sarcopterygii. Ils ont des nageoires charnues, lobées , appariées, qui sont reliées au corps par un seul os. Les nageoires des poissons à nageoires lobées diffèrent de celles de tous les autres poissons en ce que chacune est portée sur une tige charnue, en forme de lobe et écailleuse s'étendant du corps. Les nageoires pectorales et pelviennes ont des articulations ressemblant à celles des membres tétrapodes. Ces nageoires ont évolué en pattes des premiers vertébrés terrestres tétrapodes, les amphibiens . Ils possèdent également deux nageoires dorsales avec des bases séparées, contrairement à la seule nageoire dorsale des poissons à nageoires rayonnées .
Le cœlacanthe est un poisson à nageoires lobées qui existe toujours. On pense qu'il a évolué vers sa forme actuelle il y a environ 408 millions d'années, au début du Dévonien. La locomotion des coelacanthes est unique en leur genre. Pour se déplacer, les coelacanthes profitent le plus souvent des remontées ou des descentes du courant et de la dérive. Ils utilisent leurs nageoires paires pour stabiliser leur mouvement dans l'eau. Au fond de l'océan, leurs nageoires paires ne sont utilisées pour aucun type de mouvement. Les coelacanthes peuvent créer une poussée pour des démarrages rapides en utilisant leurs nageoires caudales. En raison du nombre élevé de nageoires qu'ils possèdent, les coelacanthes ont une grande maniabilité et peuvent orienter leur corps dans presque toutes les directions dans l'eau. On les a vus faire le poirier et nager le ventre en l'air. On pense que leur organe rostral aide à donner l'électroperception du cœlacanthe, ce qui facilite leur mouvement autour des obstacles.
Les dipneustes sont également des poissons vivants à nageoires lobes. Ils sont présents en Afrique ( Protopterus ), en Australie ( Neoceratodus ) et en Amérique du Sud ( Lepidosiren ).
Diversité des nageoires chez les poissons à nageoires lobes

Poisson-poumon tacheté Protopterus dolloi
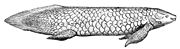
Poisson-poumon du Queensland Neoceratodus forsteri





Ray-nageoires

Les poissons à nageoires rayonnées forment une classe de poissons osseux appelés Actinopterygii. Leurs nageoires contiennent des épines ou des rayons. Une nageoire peut contenir uniquement des rayons épineux, uniquement des rayons mous ou une combinaison des deux. Si les deux sont présents, les rayons épineux sont toujours antérieurs . Les épines sont généralement raides et pointues. Les rayons sont généralement mous, flexibles, segmentés et peuvent être ramifiés. Cette segmentation des rayons est la principale différence qui les sépare des épines ; les épines peuvent être flexibles chez certaines espèces, mais elles ne seront jamais segmentées.
Les épines ont une variété d'utilisations. Chez le poisson- chat , ils sont utilisés comme forme de défense; de nombreux poissons-chats ont la capacité de verrouiller leurs épines vers l'extérieur. Les balistes utilisent également des épines pour s'enfermer dans les crevasses afin d'éviter qu'ils ne soient arrachés.
Les lépidotriches sont généralement composées d' os , mais chez les premiers ostéichthyens comme Cheirolepis , il y avait aussi de la dentine et de l'émail . Ils sont segmentés et apparaissent comme une série de disques empilés les uns sur les autres. Ils peuvent provenir d'écailles dermiques. On pense que la base génétique de la formation des rayons des nageoires est constituée de gènes codés pour la production de certaines protéines. Il a été suggéré que l'évolution du membre tétrapode des poissons à nageoires lobes est liée à la perte de ces protéines.
Diversité des nageoires chez les poissons à nageoires rayonnées
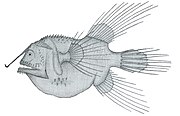
Pêcheur à la ligne Caulophryne jordani

Crêpe platax Halieutichthys aculeatus

Poisson- lune élancé Ranzania laevis

Fanfish Pteraclis carolinus

Hachette diaphane Sternoptyx diaphana

Hoplostète argenté Hoplostethus mediterraneus

Poisson-couteau Equetus lanceolatus

Pomfret atlantique Brama brama

Épave atlantique Polyprion americanus

Poisson- globe étoilé Arothron stellatus
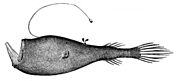
Ceratias uranoscopus d'observation des étoiles

Poromitra unicornis à tête de crête

Poisson volant tropical à deux ailes Exocoetus evolans

Benthocomète Benthocometes robustus

Queue de rat Trachonurus sulcatus

Poisson trépied Bathypterois grallator

Rameur géant Regalecus glesne

Espadon à bec court Tetrapturus angustirostris

Couteau fantôme Sternarchorhynchus oxyrhynchus

Remora Remora brachyptères

Rockskipper en pointillés bleus Blenniella periophthalmus

Bichir du Nil Polypterus bichir

Truite fardée côtière Oncorhynchus clarkii

Poisson -chat beurre africain Schilbe mystus


.gif)














.gif)
.gif)




Poissons cartilagineux


Les poissons cartilagineux forment une classe de poissons appelés Chondrichthyes. Ils ont des squelettes faits de cartilage plutôt que d' os . La classe comprend les requins , les raies et les chimères . Les squelettes d'ailerons de requin sont allongés et soutenus par des rayons mous et non segmentés appelés ceratotrichia, des filaments de protéine élastique ressemblant à la kératine cornée des cheveux et des plumes. À l'origine, les ceintures pectorales et pelviennes, qui ne contiennent aucun élément cutané, ne se connectaient pas. Dans les formes ultérieures, chaque paire de nageoires est devenue connectée ventralement au milieu lorsque les barres scapulocoracoïdes et puboischiadiques ont évolué. Chez les rayons , les nageoires pectorales sont reliées à la tête et sont très souples. L'une des principales caractéristiques présentes chez la plupart des requins est la queue hétérocerque, qui facilite la locomotion. La plupart des requins ont huit nageoires. Les requins ne peuvent s'éloigner que des objets directement devant eux car leurs nageoires ne leur permettent pas de se déplacer dans le sens de la queue en premier.
Comme pour la plupart des poissons, la queue des requins fournit une poussée, ce qui rend la vitesse et l'accélération dépendantes de la forme de la queue. La forme des nageoires caudales varie considérablement d'une espèce de requin à l'autre, en raison de leur évolution dans des environnements distincts. Les requins possèdent une nageoire caudale hétérocerque dans laquelle la partie dorsale est généralement sensiblement plus grande que la partie ventrale . En effet, la colonne vertébrale du requin s'étend dans cette partie dorsale, offrant une plus grande surface pour la fixation musculaire . Cela permet une locomotion plus efficace parmi ces poissons cartilagineux à flottabilité négative . En revanche, la plupart des poissons osseux possèdent une nageoire caudale homocerque .
Les requins tigres ont un grand lobe supérieur , ce qui permet une croisière lente et des rafales de vitesse soudaines. Le requin tigre doit être capable de se tordre et de se retourner facilement dans l'eau lors de la chasse pour soutenir son alimentation variée, tandis que le requin taupe commun , qui chasse les bancs de poissons comme le maquereau et le hareng , a un grand lobe inférieur pour l'aider à suivre le rythme de son rythme rapide. - proie nageuse. D'autres adaptations de la queue aident les requins à attraper leurs proies plus directement, comme l' utilisation par le requin -renard de son puissant lobe supérieur allongé pour étourdir les poissons et les calmars.
Aileron de requin
Selon la Humane Society International , environ 100 millions de requins sont tués chaque année pour leurs ailerons, dans un acte connu sous le nom de shark finning . Une fois les nageoires coupées, les requins mutilés sont rejetés à l'eau et laissés pour morts.
Dans certains pays d' Asie , les ailerons de requin sont un délice culinaire, comme la soupe aux ailerons de requin . Actuellement, les préoccupations internationales concernant la durabilité et le bien -être des requins ont eu un impact sur la consommation et la disponibilité de la soupe aux ailerons de requin dans le monde. Le shark finning est interdit dans de nombreux pays.
Fonctions des palmes
Poussée génératrice
Les ailerons en forme de feuille génèrent une poussée lorsqu'ils sont déplacés, la portance de l'aileron met l'eau ou l'air en mouvement et pousse l'aileron dans la direction opposée. Les animaux aquatiques obtiennent une poussée importante en déplaçant les nageoires d'avant en arrière dans l'eau. La nageoire caudale est souvent utilisée, mais certains animaux aquatiques génèrent une poussée à partir des nageoires pectorales .

Les pinnules peuvent influencer la façon dont un vortex se développe autour de la nageoire caudale.
La cavitation se produit lorsqu'une pression négative provoque la formation de bulles (cavités) dans un liquide, qui s'effondrent ensuite rapidement et violemment. Cela peut causer des dommages et une usure importants. Des dommages par cavitation peuvent se produire sur les nageoires caudales d'animaux marins puissants, tels que les dauphins et les thons. La cavitation est plus susceptible de se produire près de la surface de l'océan, où la pression de l'eau ambiante est relativement faible. Même s'ils ont le pouvoir de nager plus vite, les dauphins peuvent devoir limiter leur vitesse car l'effondrement des bulles de cavitation sur leur queue est trop douloureux. La cavitation ralentit également le thon, mais pour une raison différente. Contrairement aux dauphins, ces poissons ne sentent pas les bulles, car ils ont des nageoires osseuses sans terminaisons nerveuses. Néanmoins, ils ne peuvent pas nager plus vite car les bulles de cavitation créent un film de vapeur autour de leurs nageoires qui limite leur vitesse. Des lésions ont été trouvées sur des thons qui correspondent à des dommages de cavitation.
Les poissons scombridés (thon, maquereau et bonite) sont des nageurs particulièrement performants. Le long de la marge à l'arrière de leur corps se trouve une ligne de petites nageoires sans rayons et non rétractables, appelées pinnules . Il y a eu beaucoup de spéculations sur la fonction de ces finlets. Des recherches effectuées en 2000 et 2001 par Nauen et Lauder ont indiqué que "les nageoires ont un effet hydrodynamique sur le débit local pendant la nage régulière" et que "la nageoire la plus postérieure est orientée pour rediriger le flux dans le vortex de queue en développement, ce qui peut augmenter la poussée produite par la queue du maquereau nageur".
Les poissons utilisent plusieurs nageoires, il est donc possible qu'une nageoire donnée puisse avoir une interaction hydrodynamique avec une autre nageoire. En particulier, les nageoires immédiatement en amont de la nageoire caudale (queue) peuvent être des nageoires proches qui peuvent affecter directement la dynamique de l'écoulement au niveau de la nageoire caudale. En 2011, des chercheurs utilisant des techniques d'imagerie volumétrique ont pu générer "les premières vues tridimensionnelles instantanées des structures de sillage telles qu'elles sont produites par des poissons nageant librement". Ils ont constaté que "les battements continus de la queue entraînaient la formation d'une chaîne liée d'anneaux vortex" et que "les sillages des nageoires dorsale et anale sont rapidement entraînés par le sillage de la nageoire caudale, approximativement dans le laps de temps d'un battement de queue ultérieur".
Contrôler le mouvement
Une fois le mouvement établi, le mouvement lui-même peut être contrôlé à l'aide d'autres ailettes.


.jpg)
Les corps des poissons de récif ont souvent une forme différente de celle des poissons d'eau libre . Les poissons d'eau libre sont généralement construits pour la vitesse, profilés comme des torpilles pour minimiser la friction lorsqu'ils se déplacent dans l'eau. Les poissons de récif opèrent dans les espaces relativement confinés et les paysages sous-marins complexes des récifs coralliens . Car cette maniabilité est plus importante que la vitesse en ligne droite, c'est pourquoi les poissons des récifs coralliens ont développé des corps qui optimisent leur capacité à s'élancer et à changer de direction. Ils déjouent les prédateurs en esquivant les fissures du récif ou en jouant à cache-cache autour des têtes de corail. Les nageoires pectorales et pelviennes de nombreux poissons de récif, tels que les poissons- papillons , les demoiselles et les poissons- anges , ont évolué pour pouvoir agir comme des freins et permettre des manœuvres complexes. De nombreux poissons de récif, tels que les poissons- papillons , les demoiselles et les poissons- anges , ont des corps évolués qui sont profonds et comprimés latéralement comme une crêpe, et s'insèrent dans les fissures des rochers. Leurs nageoires pelviennes et pectorales ont évolué différemment, elles agissent donc de concert avec le corps aplati pour optimiser la maniabilité. Certains poissons, tels que le poisson-globe , le poisson - lime et le poisson -tronc , dépendent des nageoires pectorales pour nager et utilisent à peine les nageoires caudales.
la reproduction


Les poissons cartilagineux mâles (requins et raies), ainsi que les mâles de certains poissons à nageoires rayonnées vivants , ont des nageoires qui ont été modifiées pour fonctionner comme des organes intromittents , des appendices reproducteurs qui permettent la fécondation interne . Chez les poissons à nageoires rayonnées, on les appelle gonopodes ou andropodes , et chez les poissons cartilagineux, on les appelle claspers .
Les gonopodes se trouvent sur les mâles de certaines espèces des familles Anablepidae et Poeciliidae . Ce sont des nageoires anales qui ont été modifiées pour fonctionner comme des organes intromittents mobiles et sont utilisées pour imprégner les femelles de laitance pendant l'accouplement. Les troisième, quatrième et cinquième rayons de la nageoire anale du mâle forment une structure en forme de tube dans laquelle le sperme du poisson est éjecté. Lorsqu'il est prêt pour l'accouplement, le gonopode devient droit et pointe vers la femelle. Le mâle insère brièvement l'organe dans l'ouverture sexuelle de la femelle, avec des adaptations en forme de crochet qui permettent au poisson de s'agripper à la femelle pour assurer l'imprégnation. Si une femelle reste immobile et que son partenaire entre en contact avec son évent avec son gonopode, elle est fécondée. Le sperme est conservé dans l'oviducte de la femelle. Cela permet aux femelles de se féconder à tout moment sans aide supplémentaire des mâles. Chez certaines espèces, le gonopode peut mesurer la moitié de la longueur totale du corps. Parfois, la nageoire est trop longue pour être utilisée, comme dans les races "lyretail" de Xiphophorus helleri . Les femelles traitées aux hormones peuvent développer des gonopodes. Ceux-ci sont inutiles pour la reproduction.
Des organes similaires avec des caractéristiques similaires se retrouvent chez d'autres poissons, par exemple l' andropode chez l' Hemirhamphodon ou chez les Goodeidae ou le gonopode au Trias moyen † Saurichthys , le plus ancien exemple connu de viviparité chez un poisson à nageoires rayonnées.
Les agrafes se trouvent sur les mâles des poissons cartilagineux . Ce sont la partie postérieure des nageoires pelviennes qui ont également été modifiées pour fonctionner comme des organes intromittents et sont utilisées pour canaliser le sperme dans le cloaque de la femelle pendant la copulation. L'acte d'accouplement chez les requins comprend généralement l'élévation de l'un des fermoirs pour permettre à l'eau d'entrer dans un siphon à travers un orifice spécifique . Le fermoir est ensuite inséré dans le cloaque, où il s'ouvre comme un parapluie pour ancrer sa position. Le siphon commence alors à se contracter en expulsant l'eau et le sperme.
Autres fonctions
Les autres utilisations des nageoires comprennent la marche et le perchoir sur le fond marin, le glissement sur l'eau, le refroidissement de la température corporelle, l'étourdissement des proies, la parade (effrayer les prédateurs, parade nuptiale), la défense (épines venimeuses des nageoires, verrouillage entre les coraux), leurre de proies, et les structures de fixation.
Le voilier de l'Indo-Pacifique a une nageoire dorsale proéminente. Comme les scombroïdes et autres istiophoridés , ils se rationalisent en rétractant leurs nageoires dorsales dans une rainure de leur corps lorsqu'ils nagent. L'énorme nageoire dorsale, ou voile, du voilier est rétractée la plupart du temps. Les voiliers les élèvent s'ils veulent rassembler un banc de petits poissons, et aussi après des périodes de forte activité, probablement pour se rafraîchir.
_in_Waikiki_Aquarium.JPG)




Le grondin volant oriental a de grandes nageoires pectorales qu'il maintient normalement contre son corps et se dilate lorsqu'il est menacé d'effrayer les prédateurs. Malgré son nom, c'est un poisson démersal , pas un poisson volant, et utilise ses nageoires pelviennes pour marcher au fond de l'océan.
Les nageoires peuvent avoir une signification adaptative en tant qu'ornements sexuels. Pendant la parade nuptiale, le cichlidé femelle , Pelvicachromis taeniatus , affiche une grande nageoire pelvienne violette visuellement saisissante . "Les chercheurs ont découvert que les mâles préféraient clairement les femelles avec une nageoire pelvienne plus grande et que les nageoires pelviennes poussaient de manière plus disproportionnée que les autres nageoires des poissons femelles."






Évolution
Evolution des palmes paires
Il existe deux hypothèses dominantes qui ont été historiquement débattues comme modèles de l'évolution des nageoires appariées chez les poissons : la théorie de l'arc branchial et la théorie du pli latéral des nageoires. La première, communément appelée «l' hypothèse de Gegenbaur », a été posée en 1870 et propose que «les nageoires appariées sont dérivées de structures branchiales». Cela est tombé en désuétude au profit de la théorie des plis latéraux des nageoires, suggérée pour la première fois en 1877, qui propose que les nageoires appariées bourgeonnent à partir de plis latéraux longitudinaux le long de l'épiderme juste derrière les branchies. Il y a un faible soutien pour les deux hypothèses dans les archives fossiles et en embryologie. Cependant, les connaissances récentes de la structuration du développement ont incité à reconsidérer les deux théories afin de mieux élucider les origines des nageoires appariées.
Théories classiques
Le concept de Karl Gegenbaur de «l'Archipterygium» a été introduit en 1876. Il a été décrit comme un rayon branchial, ou «tige cartilagineuse jointe», qui s'étendait de l'arc branchial. Des rayons supplémentaires sont nés le long de l'arc et du rayon branchial central. Gegenbaur a suggéré un modèle d' homologie transformatrice - que toutes les nageoires et membres appariés de vertébrés étaient des transformations de l' Archipterygium . Sur la base de cette théorie, des appendices appariés tels que les nageoires pectorales et pelviennes se seraient différenciés des arcs branchiaux et auraient migré vers l'arrière. Cependant, il y a eu un soutien limité pour cette hypothèse dans les archives fossiles à la fois morphologiquement et phylogéniquement. De plus, il y avait peu ou pas de preuves d'une migration antéro-postérieure des nageoires pelviennes. De telles lacunes de la théorie de l'arc branchial ont conduit à sa disparition précoce au profit de la théorie du pli latéral des nageoires proposée par St. George Jackson Mivart , Francis Balfour et James Kingsley Thacher .
La théorie des plis latéraux des nageoires a émis l'hypothèse que des nageoires appariées se sont développées à partir de plis latéraux le long de la paroi corporelle du poisson. Tout comme la segmentation et le bourgeonnement du pli de la nageoire médiane ont donné naissance aux nageoires médianes, un mécanisme similaire de segmentation et d'allongement des bourgeons de nageoire à partir d'un pli de nageoire latéral a été proposé pour avoir donné naissance aux nageoires pectorales et pelviennes appariées. Cependant, il y avait peu de preuves d'une transition latérale du pli à la nageoire dans les archives fossiles. De plus, il a été démontré plus tard phylogéniquement que les nageoires pectorales et pelviennes proviennent d'origines évolutives et mécanistes distinctes.
Biologie du développement évolutif
Des études récentes sur l'ontogénie et l'évolution des appendices appariés ont comparé des vertébrés sans nageoires - comme les lamproies - avec des chondricthyens , le vertébré vivant le plus basal avec des nageoires appariées. En 2006, des chercheurs ont découvert que la même programmation génétique impliquée dans la segmentation et le développement des nageoires médianes se retrouvait dans le développement d'appendices appariés chez les catsharks . Bien que ces résultats ne soutiennent pas directement l'hypothèse du pli des nageoires latérales, le concept original d'un mécanisme de développement évolutif des nageoires médianes partagées reste pertinent.
Une rénovation similaire d'une ancienne théorie peut être trouvée dans la programmation du développement des arcs branchiaux chondricthyens et des appendices appariés. En 2009, des chercheurs de l' Université de Chicago ont démontré qu'il existe des mécanismes de structuration moléculaire partagés dans le développement précoce de l'arc branchial chondricthyen et des nageoires appariées. De telles découvertes ont incité à reconsidérer la théorie autrefois démystifiée de l'arc branchial.
Des nageoires aux membres
Les poissons sont les ancêtres de tous les mammifères, reptiles, oiseaux et amphibiens. En particulier, les tétrapodes terrestres (animaux à quatre pattes) ont évolué à partir de poissons et ont fait leurs premières incursions sur terre il y a 400 millions d'années. Ils utilisaient des paires de nageoires pectorales et pelviennes pour la locomotion. Les nageoires pectorales se sont développées en pattes antérieures (bras dans le cas des humains) et les nageoires pelviennes se sont développées en pattes postérieures. Une grande partie de la machinerie génétique qui construit un membre ambulant chez un tétrapode est déjà présente dans la nageoire nageuse d'un poisson.
Aristote reconnut la distinction entre les structures analogues et homologues , et fit la comparaison prophétique suivante : « Les oiseaux ressemblent en quelque sorte aux poissons. Car les oiseaux ont leurs ailes dans la partie supérieure de leur corps et les poissons ont deux nageoires dans la partie antérieure de leur corps. Les oiseaux ont des pieds sur leur partie inférieure et la plupart des poissons ont une deuxième paire de nageoires dans leur partie inférieure et près de leurs nageoires avant."
– Aristote, De incessu animalium


En 2011, des chercheurs de l'Université Monash en Australie ont utilisé des poissons-poumons primitifs mais encore vivants " pour retracer l'évolution des muscles de la nageoire pelvienne afin de découvrir comment les membres postérieurs porteurs des tétrapodes ont évolué". Des recherches plus poussées à l'Université de Chicago ont révélé que les poissons-poumons marchant sur le fond avaient déjà développé les caractéristiques des allures de marche des tétrapodes terrestres.
Dans un exemple classique d' évolution convergente , les membres pectoraux des ptérosaures , des oiseaux et des chauves -souris ont ensuite évolué le long de voies indépendantes en ailes volantes. Même avec des ailes volantes, il existe de nombreuses similitudes avec les jambes qui marchent, et les aspects fondamentaux de l'empreinte génétique de la nageoire pectorale ont été conservés.
Les premiers mammifères sont apparus au Permien (entre 298,9 et 252,17 millions d'années). Plusieurs groupes de ces mammifères ont commencé à retourner à la mer, dont les cétacés (baleines, dauphins et marsouins). Une analyse ADN récente suggère que les cétacés ont évolué à partir des ongulés à doigts pairs et qu'ils partagent un ancêtre commun avec l' hippopotame . Il y a environ 23 millions d'années, un autre groupe de mammifères terrestres ressemblant à des ours a commencé à retourner dans la mer. C'étaient les sceaux . Ce qui était devenu des membres ambulants chez les cétacés et les phoques a évolué indépendamment vers de nouvelles formes de nageoires nageuses. Les membres antérieurs sont devenus des nageoires , tandis que les membres postérieurs ont été soit perdus (cétacés), soit également modifiés en nageoires (pinnipèdes). Chez les cétacés, la queue a gagné deux nageoires à la fin, appelées douves . Les queues de poisson sont généralement verticales et se déplacent d'un côté à l'autre. Les douves des cétacés sont horizontales et se déplacent de haut en bas, car les épines des cétacés se plient de la même manière que chez les autres mammifères.

Les ichtyosaures sont d'anciens reptiles qui ressemblaient à des dauphins. Ils sont apparus pour la première fois il y a environ 245 millions d'années et ont disparu il y a environ 90 millions d'années.
"Ce reptile marin avec des ancêtres terrestres a convergé si fortement sur les poissons qu'il a en fait développé une nageoire dorsale et une nageoire caudale pour une locomotion aquatique améliorée. Ces structures sont d'autant plus remarquables qu'elles ont évolué à partir de rien - le reptile terrestre ancestral n'avait pas de bosse sur son dos ou sa lame sur sa queue pour servir de précurseur."
Le biologiste Stephen Jay Gould a déclaré que l'ichtyosaure était son exemple préféré d' évolution convergente .
Des nageoires ou des nageoires de formes variées et à divers endroits (membres, corps, queue) ont également évolué dans un certain nombre d'autres groupes de tétrapodes, y compris les oiseaux plongeurs tels que les pingouins (modifiés à partir des ailes), les tortues de mer (membres antérieurs modifiés en nageoires), les mosasaures (membres modifiés en nageoires) et les serpents de mer (nageoire caudale élargie verticalement et aplatie).
Ailerons robotiques

| Vidéo externe | |
|---|---|
|
| |
|
| |
|
| |
|
| |
|
|
L'utilisation de nageoires pour la propulsion des animaux aquatiques peut être remarquablement efficace. Il a été calculé que certains poissons peuvent atteindre une efficacité de propulsion supérieure à 90 %. Les poissons peuvent accélérer et manœuvrer beaucoup plus efficacement que les bateaux ou les sous- marins , et produire moins de perturbations de l'eau et de bruit. Cela a conduit à des études biomimétiques de robots sous-marins qui tentent d'imiter la locomotion des animaux aquatiques. Un exemple est le Robot Tuna construit par l' Institute of Field Robotics , pour analyser et modéliser mathématiquement le mouvement thunniforme . En 2005, le Sea Life London Aquarium a présenté trois poissons robotiques créés par le département d'informatique de l' Université d'Essex . Les poissons ont été conçus pour être autonomes, nager et éviter les obstacles comme de vrais poissons. Leur créateur a affirmé qu'il essayait de combiner "la vitesse du thon, l'accélération d'un brochet et les compétences de navigation d'une anguille".
L' AquaPenguin , développé par Festo en Allemagne, copie la forme profilée et la propulsion par les nageoires avant des pingouins . Festo a également développé AquaRay , AquaJelly et AiraCuda , émulant respectivement la locomotion des raies manta, des méduses et des barracudas.
En 2004, Hugh Herr du MIT a prototypé un poisson robotique biomécatronique avec un actionneur vivant en transplantant chirurgicalement les muscles des cuisses de grenouille au robot, puis en faisant nager le robot en pulsant les fibres musculaires avec de l'électricité.
Les poissons robotisés offrent certains avantages en matière de recherche, tels que la possibilité d'examiner une partie individuelle d'une conception de poisson isolément du reste du poisson. Cependant, cela risque de trop simplifier la biologie, de sorte que des aspects clés de la conception de l'animal sont négligés. Les poissons robots permettent également aux chercheurs de faire varier un seul paramètre, comme la flexibilité ou un contrôle de mouvement spécifique. Les chercheurs peuvent mesurer directement les forces, ce qui n'est pas facile à faire chez les poissons vivants. "Les dispositifs robotiques facilitent également les études cinématiques tridimensionnelles et les analyses hydrodynamiques corrélées, car l'emplacement de la surface locomotrice peut être connu avec précision. Et, les composants individuels d'un mouvement naturel (tels que la course vers l'avant par rapport à la course d'un appendice battant) peuvent être programmés séparément, ce qui est certainement difficile à réaliser lorsque l'on travaille avec un animal vivant."
Voir également
- Nageoire céphalopode
- Locomotion des nageoires et des nageoires
- Locomotion des poissons
- Polydactylie chez les premiers tétrapodes
- RobotTuna
- Soupe aux ailerons de requin
- Compromis pour la locomotion dans l'air et dans l'eau
- Locomotion ondulatoire
Références
Citations
Bibliographie
-
En ligneHamlett, William C. (1999). Requins, patins et raies: la biologie des poissons élasmobranches (1 éd.). p 56: La Presse de l'Université Johns Hopkins. ISBN 978-0-8018-6048-5.
{{cite book}}: Maint CS1: emplacement ( lien )
Lectures complémentaires
- Hall, Brian K (2007) Fins into Limbs: Evolution, Development, and Transformation University of Chicago Press. ISBN 9780226313375 .
- Helfman G, Collette BB, Facey DE et Bowen BW (2009) "Morphologie fonctionnelle de la locomotion et de l'alimentation" Chapitre 8, pp. 101–116. Dans : La diversité des poissons : Biologie , John Wiley & Sons. ISBN 9781444311907 .
- Lauder, GV ; Nauen, JC; Drucker, EG (2002). "Hydrodynamique expérimentale et évolution: fonction des nageoires médianes chez les poissons à nageoires rayonnées" . Intégr. Comp. Biol . 42 (5): 1009-1017. doi : 10.1093/icb/42.5.1009 . PMID 21680382 .
- Lauder, GV ; Drucker, EG (2004). "Morphologie et hydrodynamique expérimentale des surfaces de contrôle des nageoires de poisson" (PDF) . Journal de génie océanique . 29 (3): 556–571. Bibcode : 2004IJOE...29..556L . doi : 10.1109/joe.2004.833219 . S2CID 36207755 .
Liens externes
- Homologie des lépidotrichies des nageoires chez les poissons ostéichtyens
- L'aileron du poisson Earthlife Web
- Les poissons robots peuvent-ils trouver la pollution ? HowStuffWorks . Consulté le 30 janvier 2012.