
Récepteur du peptide formyl - Formyl peptide receptor
| récepteur du formyl peptide 1 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiants | |||||||
| symbole | FPR1 | ||||||
| Alt. symboles | FPR; FMLP | ||||||
| gène NCBI | 2357 | ||||||
| HGNC | 3826 | ||||||
| OMIM | 136537 | ||||||
| RéfSeq | NM_002029 | ||||||
| UniProt | P21462 | ||||||
| Autre informations | |||||||
| Lieu | Chr. 19 q13.41 | ||||||
| |||||||
| récepteur du formyl peptide 2 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiants | |||||||
| symbole | FPR2 | ||||||
| Alt. symboles | ALXR, FMLPX, FPR2/ALX, FPR2A, FPRH1, FPRL1, HM63, LXA4R, RFP | ||||||
| gène NCBI | 2358 | ||||||
| HGNC | 3827 | ||||||
| OMIM | 136538 | ||||||
| RéfSeq | NM_001462 | ||||||
| UniProt | P25090 | ||||||
| Autre informations | |||||||
| Lieu | Chr. 19 q13.3-13.4 | ||||||
| |||||||
| récepteur de formyl peptide 3 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiants | |||||||
| symbole | FPR3 | ||||||
| Alt. symboles | FPRH2, FPRL2, FMLPY | ||||||
| gène NCBI | 2359 | ||||||
| HGNC | 3828 | ||||||
| OMIM | 136539 | ||||||
| RéfSeq | NM_002030 | ||||||
| UniProt | P25089 | ||||||
| Autre informations | |||||||
| Lieu | Chr. 19 q13.3-13.4 | ||||||
| |||||||
Les récepteurs formyl peptide s ( FPR ) appartiennent à une classe de récepteurs couplés aux protéines G impliqués dans la chimiotaxie . Chez l'homme, il existe trois isoformes du récepteur du formyl peptide, chacune codée par un gène distinct nommé FPR1 , FPR2 et FPR3 . Ces récepteurs ont été identifiés à l'origine par leur capacité à se lier à des peptides N-formyl tels que la N- formylméthionine produite par la dégradation de cellules bactériennes ou hôtes. Par conséquent, les récepteurs du formyl peptide sont impliqués dans la médiation de la réponse des cellules immunitaires à l'infection. Ces récepteurs peuvent également agir pour supprimer le système immunitaire dans certaines conditions. La relation phylogénétique étroite de la signalisation dans la chimiotaxie et l'olfaction a été récemment prouvée par la détection de protéines de type récepteur de formyl peptide en tant que famille distincte de chimiocapteurs d'organes voméronasaux chez la souris.
Le FPR est maintenant correctement accepté sous le nom de FPR1 par l'Union internationale de pharmacologie fondamentale et clinique.
Découverte
Des études menées dans les années 1970 ont révélé qu'une série d' oligopeptides contenant de la N-formylméthionine , dont le membre le plus puissant et le plus connu de cette série, la N-formylméthionine-leucyl-phénylalanine (fMLF ou fMet-Leu-Phe), stimulaient le lapin et l'humain. les neutrophiles par un mécanisme apparent dépendant du récepteur pour migrer selon un schéma directionnel dans les tests de chimiotaxie classiques en laboratoire . Étant donné que ces oligopeptides ont été produits par des bactéries ou des analogues synthétiques de tels produits, il a été suggéré que les oligopeptides N-formyl sont d'importants facteurs chimiotiques et que leurs récepteurs sont d'importants récepteurs de facteurs chimiotactiques qui agissent respectivement comme éléments de signalisation et de reconnaissance de signaux pour initier des réponses inflammatoires dans afin de se défendre contre l'invasion bactérienne. D'autres études ont défini un récepteur pour les oligopeptides N-formyl, le récepteur du peptide formyl (FPR), ainsi nommé en fonction de sa capacité à se lier et à être activé par les oligopeptides. Deux récepteurs ont ensuite été découverts et nommés FPR1 et FPR2 sur la base de la similitude de la séquence d'acides aminés prédite de leurs gènes avec celle de FPR plutôt que sur une quelconque capacité à se lier ou à être activé par les oligopeptides formyl. Ces trois récepteurs ont été renommés FPR1, FPR2 et FPR3 et se sont avérés avoir des spécificités très différentes pour les oligopeptides formyls et des fonctions très différentes qui incluent l'initiation de réponses inflammatoires aux peptides N-formyl libérés non seulement par les bactéries mais également une multiplicité d'éléments libérés. par les tissus de l'hôte ; amortir et résoudre les réponses inflammatoires; et peut-être contribuer au développement de certains cancers neurologiques et d'un éventail de maladies neurologiques Maladies à base d' amyloïde .
La structure et la fonction
Le récepteur formyl peptide (FPR) appartient à la classe des récepteurs possédant sept domaines transmembranaires hydrophobes. La conformation du FPR est stabilisée par plusieurs interactions. Ceux-ci incluent la formation potentielle de ponts salins entre Arg84-Arg205, Lys85-Arg205 et Lys85-Asp284 qui aident à déterminer la structure tridimensionnelle des domaines transmembranaires, ainsi que les résidus chargés positivement (Arg, Lys) qui interagissent avec les phosphates chargés négativement. De plus, le résidu Arg163 peut interagir avec la poche de liaison au ligand de la deuxième boucle extracellulaire du FPR.
En ce qui concerne la liaison du peptide formyle Met-Leu-Phe, il existe des interactions potentielles supplémentaires qui incluent des interactions de liaison hydrogène entre Arg84 et Lys85 de la première boucle extracellulaire et le groupe N-formyle du ligand ainsi que le squelette peptidique du formyle. Met-Leu-Phe qui peut former des interactions similaires. La fraction formyl-Met du ligand s'est avérée former des ponts disulfure avec les résidus Cys, et une interaction avec Arg163 a également été démontrée. (Il est important de mentionner que certaines interactions qui stabilisent la conformation du récepteur peuvent également influencer la liaison au ligand.) Certains oligopeptides ont également été décrits comme des constituants caractéristiques liés à l'Asn-s de la partie N-terminale extracellulaire et à la poche de liaison du ligand de la deuxième boucle extracellulaire. Ces composants peuvent également déterminer ou rendre plus spécifique l'interaction ligand-récepteur.

Diagramme schématique du récepteur du formyl peptide 1. Les hélices transmembranaires du récepteur sont représentées par des cylindres bleu-vert tandis que la membrane cellulaire dans laquelle le récepteur est noyé est représentée en jaune. La face extracellulaire de la membrane cellulaire est en haut tandis que la face intracellulaire ( cytoplasmique ) est en bas. Les boucles extracellulaires du FPR responsables de la liaison N-for-Met-Leu-Phe (Nfor-MLF) sont indiquées en rouge.
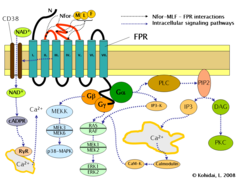
Voies de signalisation du récepteur du formyl peptide (FPR).


Voies de signalisation
L'induction de la FPR déclenche de multiples changements dans les cellules eucaryotes , y compris le réarrangement du cytosquelette qui à son tour facilite la migration cellulaire et la synthèse de chimiokines . Les voies réglementées par le FPR comprennent :
- Activation dépendante de la protéine G de la phospholipase C (PLC) qui entraîne la décomposition du phospholipide constitutif de la membrane, le phosphatidylinositol (4,5)-bisphosphate (PIP2) en inositol (1,4,5)-triphosphate (IP3) et en diacyl glycérol ( DAG). IP3 est l'un des inducteurs les plus efficaces de l' augmentation du Ca 2+ à partir des pools cytoplasmiques et de l'extérieur de la cellule via l'ouverture des canaux Ca 2+ . Le DAG est à son tour un inducteur de la protéine kinase C (PKC).
- Activation de la petite GTPase régulatrice, RAS . Le RAS actif peut à son tour activer RAF , une kinase Ser/Thr. Dans l'étape suivante, les protéines kinases activées par les mitogènes (MAP kinases) sont activées. (Également connues sous le nom de kinases régulées par le signal extracellulaire - ERK ou MAP/ERK kinase (MEK)). À la suite de la dernière étape, ERK1 et ERK2 sont activés. Les formes phosphorylées des ERK peuvent continuer la cascade en déclenchant l'activation de plus de kinases interagissantes, ce qui entraîne une altération de l'activité transcriptionnelle dans le noyau.
- La liaison du ligand au FPR peut également induire l'activation de CD38 , une ectoenzyme de la membrane de surface. À la suite de l'activation, les molécules NAD + entreront dans le cytoplasme. Le NAD + est converti en ADP ribose cyclique (cADPR), un second messager qui interagit avec les récepteurs de la ryanodine (RyR) à la surface du réticulum endoplasmique rugueux . Le résultat global du processus est une augmentation des niveaux de Ca 2+ cytoplasmique via la voie directe décrite ci-dessus et également via des voies indirectes telles que l'ouverture de canaux Ca 2+ dans la membrane cellulaire. L'augmentation soutenue de Ca 2+ est nécessaire pour la migration dirigée des cellules.
Voir également
Les références
Liens externes
- "Récepteurs formylpeptides" . Base de données IUPHAR des récepteurs et canaux ioniques . Union internationale de pharmacologie fondamentale et clinique.
- Formyl + peptide + récepteur à la National Library of Medicine Medical Subject Headings (MeSH) des États-Unis