
Radiodonta - Radiodonta
| Radiodonta |
|
|---|---|

|
|
| Amplectobelua symbrachiata , Anomalocaris canadensis , Aegirocassis benmoulai , Peytoia nathorsti , Lyrarapax unguispinus et Hurdia victoria | |
| Classification scientifique | |
| Royaume: | |
| Phylum: | |
| Classe: | |
| Commande: |
† Radiodonta
Collins , 1996 |
| Familles | |
Radiodonta est un ordre d' arthropodes de groupes souches qui a connu un succès dans le monde entier pendant la période cambrienne et qui comprenait les premiers grands prédateurs connus. Ils peuvent être appelés radiodontes , radiodontains , radiodontidés , anomalocarides ou anomalocaridides , bien que ces derniers se réfèrent à l'origine à la famille des Anomalocarididae , qui comprenait auparavant toutes, mais récemment seulement quelques espèces de cet ordre. Certaines des espèces les plus connues de radiodontes sont les taxons cambriens Anomalocaris canadensis , Hurdia victoria , Peytoia nathorsti et Amplectobelua symbrachiata , l' Ordovicien Aegirocassis benmoulai et le Dévonien Schinderhannes bartelsi .
Étymologie
Le nom Radiodonta (latin pour rayon "parlait d'une roue" et grec pour odoús "dent") se réfère à la disposition radiale des plaques dentaires (cône oral) entourant la bouche, bien que ces caractéristiques soient suggérées pour être absentes dans certaines espèces de radiodontes.
Définition
Le diagnostic initial de l'ordre Radiodonta en 1996 est le suivant:
Les radiodontidés sont des arthropodes allongés bilatéralement symétriques avec une cuticule non minéralisée généralement plus robuste dans les mâchoires et les griffes. Le corps est subdivisé en deux tagmata , un peu comme le prosoma et l' opisthosoma des arthropodes chélicérés . En règle générale, la partie antérieure ne présente aucune segmentation externe, porte une paire de griffes pré-orales, une paire d'yeux proéminents et des mâchoires ventrales avec des dents rayonnantes. Certaines formes ont des rangées de dents supplémentaires et trois ou quatre paires de membres gnathobasiques postoraux . Le tronc est métamérique , généralement avec environ 13 segments développant latéralement des lobes imbriqués pour la nage et des branchies pour la respiration, et peut se terminer par une queue en trois parties proéminente. Certaines formes ont des membres du tronc gnathobasiques.
En 2014, le clade Radiodonta a été défini phylogénétiquement comme un clade incluant tout taxon plus proche d' Anomalocaris canadensis que Paralithodes camtschaticus . En 2019, il a été redéfini morphologiquement en tant que complexe de carapace de tête d'animal avec des éléments centraux (H-) et latéraux (P-); excroissances (endites) des appendices frontaux portant des épines auxiliaires; et des lambeaux antérieurs réduits ou des bandes de lamelles (lames sétales) et un fort effilement du corps de l'avant vers l'arrière.

La description

La plupart des radiodontes sont significativement grands par rapport à d'autres animaux cambriens , avec une longueur de corps généralement comprise entre 30 et 50 centimètres. Le plus grand radiodonte décrit est l' Ordovicien Aegirocassis benmoulai , qui peut avoir atteint 2 mètres de long. Le Lyrarapax de 8 centimètres est l'un des plus petits radiodontes, les juvéniles de L. unguispinus ne mesurant qu'environ 18 millimètres.
Le corps d'un radiodont peut être divisé en deux régions: la tête et le tronc. La tête est composée d'un seul segment du corps connu sous le nom de somite oculaire, recouvert de sclérites (complexe de carapace de la tête), avec des appendices frontaux arthropodisés, des pièces buccales ventrales (cône oral) et des yeux composés pédonculés . Alors que le tronc effilé est composé de plusieurs segments de corps, chacun associé à des paires de volets et de structures en forme de branchies (lames de sétal).
Appendice frontal




Les structures antérieures de la tête sont une paire d'appendices frontaux. Ils peuvent être appelés `` griffes '', `` appendices de préhension '', `` appendices d'alimentation '' ou `` grands appendices '' dans les études précédentes, mais récemment, ces derniers n'étaient plus utilisés car les appendices frontaux étaient considérés comme non homologues aux grands appendices mégacheiran . Ils sont sclérisés et segmentés ( arthropodisés ), portant des épines ventrales (endites) sur la plupart de leurs unités segmentaires (podomères), et les endites peuvent porter des rangées supplémentaires d'épines (épines auxiliaires) sur leurs marges antérieures et postérieures. L'appendice frontal pourrait être divisé en deux régions: la tige («pédoncule» dans les études précédentes) et la région articulée distale. La région triangulaire couverte par la cuticule molle (membrane arthrodiale), peut se produire entre la face ventrale de chaque podomère et offrir une flexibilité. Leur origine pré-oculaire et protocérébrale suggèrent qu'ils sont homologues aux antennes primaires d' Onychophora et le labrum d' Euarthropoda (tous sont issus de somite oculaire), et non homologues avec les chélicères de Chelicerata ni les antennes et grands appendices d'autres arthropodes, qui sont deutocérébral (asore du somite post-oculaire 1). Étant donné que la morphologie des appendices frontaux, en particulier ceux des endites, diffèrent toujours entre les espèces, ils sont l'un des caractères les plus importants dans l'identification des espèces. En fait, de nombreux radiodontes ne sont connus qu'à partir d'une poignée d'appendices frontaux fossilisés.

Appendices frontaux d' Anomalocarididae , d' Amplectobeluidae et éventuellement d'espèces apparentées.

Appendices frontaux des Tamisiocarididae (= " Cetiocaridae ").

Appendices frontaux des Hurdiidae .



Cône oral


La bouche est sur la face ventrale de la tête, derrière le point d'attache des appendices frontaux et est entourée d'un anneau de plaques dentaires, formant la partie buccale appelée cône oral («mâchoires» dans les études précédentes). 3 ou 4 plaques dentaires peuvent grossir, donnant au cône buccal un aspect triradial (par exemple Anomalocaris ) ou tétraradial (par exemple Hurdiidae , Lyrarapax ). Le bord intérieur des plaques dentaires a des pointes tournées vers l'ouverture buccale. Des rangées supplémentaires de plaques dentaires internes peuvent apparaître dans certains genres hurdiid . La reconstruction détaillée des cônes oraux amplectobéluides est spéculative, mais ils ne présentaient peut-être pas une disposition radiale typique.
Sclérites de la tête, yeux et tronc




Trois sclérites de la tête (complexe carapace de la tête) formés par un élément H central (sclérite antérieure ou têtière) et une paire d'éléments P (sclérites latéraux), ils recouvrent respectivement la surface dorsale et latéroventale de la tête de l'animal. Les éléments P peuvent se connecter les uns aux autres ainsi qu'à l'élément H par une extension antérieure étroite (col de l'élément P ou «bec»). Le complexe de la carapace de la tête est petit et ovoïde chez les Anomalocarididae et Amplectobeluidae , mais souvent élargi chez les Hurdiidae . La tête portait deux yeux composés pédonculés , qui peuvent avoir une mobilité, et situés entre les espaces formés par les régions postérieures de l'élément H et de l'élément P.


Contrairement au diagnostic initial, la division des segments corporels (limites segmentaires) peut être visible de l'extérieur et aucun membre connu de Radiodonta (à l'exception du radiodont putatif Cucumericrus ) n'est connu pour avoir des appendices pédiformes du tronc (pattes). Le tronc a de nombreux segments corporels ( somites ), se rétrécissant de l'avant vers l'arrière, avec les 3 ou 4 segments antérieurs considérablement rétrécis dans une région du cou.

Les appendices du tronc étaient des lambeaux de corps en forme de nageoire (`` lambeaux latéraux '' ou `` lobes '' dans certaines études), généralement une paire de lambeaux ventral par segment corporel, chacun légèrement chevauché sur celui qui est le plus antérieur, mais supplémentaire, non chevauchante un ensemble de petits lambeaux dorsaux peut apparaître chez certaines espèces de Hurdiid . Les lambeaux peuvent avoir de nombreuses structures en forme de veine appelées rayons de renforcement. Les lambeaux de la région du cou, appelés lambeaux antérieurs ou lambeaux cervicaux, sont considérablement réduits. Chez certaines espèces, des appendices d'alimentation en forme de mâchoire appelés structures de type gnathobase (GLS) provenaient de chacune de la base de leurs rabats de cou réduits. De nombreuses extensions allongées en forme de lame (lames lancéolées) disposées en rangée, formant des bandes de structure en forme de branchies appelées lames ou lamelles en sétal, couvraient la surface dorsale de chaque segment du corps. Au moins chez Aegirocassis , chaque lame lancéolée est couverte de rides.
Les lambeaux ventraux peuvent être homologues de l'endopode des membres birames des euarthropodes et des membres lobopodes (lobopodes) des lobopodes branchus , et les lambeaux dorsaux et les lames de la sétale peuvent être homologues de l'exopode et des lambeaux dorsaux branchiaux des anciens taxons. Le tronc peut se terminer par 1 à 3 paires d'éventail de queue, deux longs furcae, une structure terminale allongée ou une pointe émoussée sans traits.

Aegirocassis , Peytoia et Hurdia . Troisgenres hurdiid qui portaient des lambeaux dorsaux et un complexe de carapace de la tête élargi.

Anomalocaris canadensis , unradiodonte Anomalocaridid avec un petit complexe de carapace de tête ovoïde, 3 paires d'éventail de queue et un cordier terminal.

Amplectobelua symbrachiata . Un radiodont Amplectobeluid avec des structures de type gnathobase et deux furcae.



Structures internes

Des traces de muscles , de système digestif et de système nerveux ont été décrites à partir de certains fossiles de radiodontes. Paires de muscles bien développés reliés aux lambeaux ventraux, situés au niveau des cavités latérales de chaque segment du corps. Entre les muscles latéraux se trouve un système digestif sophistiqué, formé par l'élargissement de l'intestin antérieur et de l'intestin postérieur, tous deux reliés par un intestin moyen étroit associé à 6 paires de divercules intestinaux (glandes digestives). Comparé au cerveau à trois segments de l' euarthropode et au cerveau à deux segments de l' onychophorane , le cerveau du radiodonte est composé d'un seul segment cérébral provenant du somite oculaire, le protocerebrum. Les nerfs des appendices frontaux et des yeux composés sont issus respectivement des régions antérieure et latérale du cerveau. En arrière du cerveau se trouvait une paire de cordons nerveux ventraux apparemment non fusionnés qui traversaient la région du cou de l'animal.
Paléoécologie
Physiologie
Les radiodontes ont été interprétés comme des animaux nektoniques ou nektobenthiques , leur morphologie suggérant un mode de vie actif de natation. Les volets ventraux musclés qui se chevauchent peuvent avoir propulsé l'animal dans l'eau, peut-être en se déplaçant dans une formation en forme de vague ressemblant à des raies modernes et à la seiche . Des paires de volets dorsaux, qui constituent un éventail de queue chez certaines espèces, peuvent aider à diriger et / ou à stabiliser l'animal pendant la locomotion. Chez Anomalocaris , la morphologie de l'éventail caudal suggère même qu'il pourrait changer rapidement sa direction de nage efficacement. Des bandes de lames sétales avec des lames lancéolées plissées peuvent augmenter la surface, suggérant qu'il s'agissait de branchies , assurant la fonction respiratoire de l'animal . L'abondance des restes de structures sclérosées telles que les appendices frontaux désarticulés et le complexe de carapace de la tête, suggèrent que des événements de mue massive peuvent s'être produits chez les radiodontes, un comportement qui a également été signalé chez certains autres arthropodes cambriens tels que les trilobites .
Régime

Les radiodontes avaient diverses stratégies d'alimentation, qui pouvaient être classées comme prédateurs rapaces , tamiseuses de sédiments ou filtreurs en suspension . Par exemple, des prédateurs rapaces comme Anomalocaris et Amplectobeluids auraient pu attraper des proies agiles en utilisant leurs appendices frontaux rapaces, ces derniers portaient même un endite robuste pour tenir des proies comme une pince . Avec le plus petit complexe de carapace de la tête et la grande surface des membranes arthrodiales, les appendices frontaux de ces taxons avaient une plus grande flexibilité. Les appendices frontaux robustes des tamis à sédiments comme Hurdia et Peytoia ont des endites dentelées avec une courbure mésiale, qui pourraient former un piège en forme de panier pour ratisser les sédiments et diriger les aliments vers le cône oral bien développé. Certaines espèces peuvent avoir d'autres caractéristiques très spécialisées pour un mode de vie de tamisage des sédiments, comme le Cambroraster avec son élément H en forme de dôme semblable à la carapace d'un crabe fer à cheval . Les endites des appendices frontaux des suspensions / filtreurs comme Tamisiocaris et Aegirocassis ont des épines auxiliaires flexibles et densément compactées , qui pourraient filtrer les composants organiques tels que le mésozooplancton et le phytoplancton jusqu'à 0,5 mm. Appendices frontaux des Caryosyntrips , qui sont inhabituels pour les radiodontes car ils ont la direction des surfaces portant des endites opposées et peuvent avoir été capables de manipuler et d'écraser des proies dans un mouvement de tranchage ou de saisie semblable à des ciseaux.


Peytoia nathorsti , un radiodonte hurdiid tamisant les sédiments avec de gros appendices frontaux, étroitement associé à un cône oral tétraradial

Appendice frontal de la suspension d'alimentation radiodont Tamisiocaris borealis , montrant endite allongée avec des épines auxiliaires densément tassées.
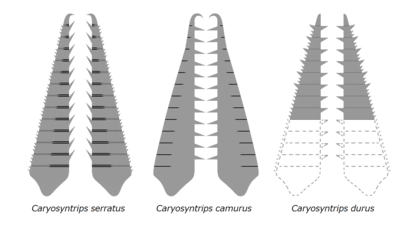
Appendices frontaux appariés de Caryosyntrips , montrant une direction opposée.

Les cônes oraux des radiodontes peuvent être capables de réaliser une aspiration et / ou une capacité de morsure. Parallèlement aux appendices frontaux, la différenciation des cônes oraux entre les genres suggère également des préférences de régime alimentaire différent. Par exemple, le cône oral triradial d' Anomalocaris avec des plaques dentaires irrégulières et tuberculées et une ouverture plus petite peut être adapté aux proies petites et actives; tandis que les cônes oraux tétraradiaux rigides de Hurdia et Cambroraster avec une ouverture plus grande et des plaques dentaires supplémentaires peuvent être capables de consommer un aliment plus gros.
Classification
Affinités taxomomiques
|
||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogénie résumée entre Radiodonta et d'autres taxons Ecdysozoan. |




La plupart des analyses suggèrent que les radiodontes sont des arthropodes de groupes souches et soeurs des deutéropodes , un clade comprenant la tige supérieure (par exemple les fuxianhuiidés et les arthropodes bivalves) et les Euarthropodes de la couronne (par exemple Artiopoda , Chelicerata et Mandibulata ). Cette interprétation est étayée par de nombreuses caractéristiques de type arthropode trouvées sur les radiodontes, telles que les yeux composés , les glandes digestives, l'arthropodisation (sur les appendices frontaux), les appendices du tronc se formant par des éléments dorsaux et ventraux (précurseur des appendices arthropodes biramus), l' intestin antérieur cuticularisé et l'intestin postérieur, ainsi que la sclérite antérieure protocérébrale. La région du cou resserrée avec les structures appendiculaires d'alimentation de certains radiodontes peut également éclairer l'origine de la tête d'arthropode sophistiquée, qui est formée par la fusion de plusieurs segments du corps antérieur.

Les 'branchies lobopodians ' Opabinia ( en haut), Pambdelurion (en bas à gauche) et Kerygmachela (en bas à droite).

Les lobopodes sibériides Siberion (en haut à gauche), Megadictyon (en bas au centre) et Jianshanopodia (en haut à droite).


Les taxons juste à la base de la branche des radiodontes et des euarthropodes sont Pambdelurion , Kerygmachela et Opabinia , trois genres de dinocaridides de type radiodont généralement appelés «lobopodiens branchiaux». Ils ont des lambeaux corporels, des glandes digestives et des appendices frontaux spécialisés comme le radiodonte, mais les appendices frontaux ne sont pas arthropodisés et ils portent un lobopode sous chacun de leurs lambeaux. Opabinia est davantage dérivée en ayant des yeux pédonculés de type radiodonte, un éventail de queue, des lames de setal et même une ouverture de la bouche postérieure de type euarthropode. Les taxons, même basaux aux «lobopodiens branchiaux», sont des sibériides comme Megadictyon et Jianshanopodia , un groupe de lobopodiens portant des appendices frontaux robustes et des glandes digestives, mais pas de lambeaux corporels. De telles formes intermédiaires entre le lobopode et le radiodonte / euarthropode suggèrent que les arthropodes du groupe total sont issus d'un grade de lobopode paraphylétique , aux côtés des deux autres panarthropodes phyla Tardigrada et Onychophora existants .


Des études antérieures peuvent suggérer les radiodontes comme un groupe autre que les arthropodes souches, tels que les vers cycloneuralia n soumis à une convergence avec les arthropodes (sur la base des pièces buccales radiales de type cycloneuralien); la tige chélicère aux côtés de megacheira ns aka grands arthropodes d'appendice (basé sur la similitude entre les appendices frontaux de radiodont, les grands appendices mégacheiran et les chélicères ); ou Schinderhannes bartelsi , qui s'est résolu comme un radiodonte hurdiid dans des analyses récentes, comme une espèce plus étroitement liée aux euarthropodes que d'autres radiodontes (sur la base de certaines caractéristiques putatives de type euarthropode trouvées sur Schinderhannes ). Cependant, ni chacun d'eux n'a été étayé par des enquêtes ultérieures. Les pièces buccales radiales ne sont pas exclusives aux cycloneuraliens et présentent plus probablement le résultat d'une évolution convergente ou d'une plésimorphie ecdysozoa n , puisqu'elles ont également été trouvées dans des panarthropodes tels que le tardigrade et certains lobopodes ; les grands appendices du mégacheiran étaient considérés comme deutocérébraux, qui ne sont pas homologues aux appendices frontaux protocérébraux du radiodont; Les caractères putatifs d'euarthropodes trouvés sur le seul fossile de Schinderhannes sont discutables et peuvent présenter d'autres structures de type radiodont.
Interrelations
Traditionnellement, Radiodonta comprenait tous les taxons de la classe Dinocaridida , et les taxons actuellement inclus dans Radiodonta ont été placés dans une famille, Anomalocarididae , d'où le nom commun `` anomalocaridid '' et il était encore parfois utilisé pour désigner l'ordre entier même après reclassification. Récemment, les genres basaux de dinocaridides Pambdelurion , Kerygmachela et Opabinia ont été placés à l'extérieur de Radiondonta, et la plupart des espèces de radiodont ont été reclassées dans trois nouvelles familles: Amplectobeluidae , Tamisiocarididae (anciennement Cetiocaridae ) et Hurdiidae . En incluant les Anomalocarididae, les quatre familles récentes de radiodontes peuvent former le clade Anomalocarida .

La description originale de l'ordre Radiodonta comprenait Anomalocaris , Laggania (plus tard connu sous le nom de Peytoia ), Hurdia , Proboscicaris , Amplectobelua , Cucumericrus et Parapeytoia . Cependant, Proboscicaris est maintenant considéré comme un synonyme plus récent de Hurdia , et Parapeytoia est considéré comme un mégacheiran. La position du Cucumericrus au sein de Radiodonta n'est pas claire, car il a été soit non sélectionné par analyse phylogénétique, soit résolu dans une polytomie avec Radiodonta et Euarthropoda .


Sous Radiodonta, Caryosyntrips est le genre le plus basal aux côtés de Cucumericrus (si inclus). Le genre Anomalocaris s'est toujours avéré non monophylétique , généralement avec Anomalocaris kunmingensis et Anomalocaris briggsi résolu en tant que membre des Amplectobeluidae et Tamisiocarididae respectivement. L'interrelation des Amplectobeluidae est incertaine, car les affinités amplectobeluidiques de Lyrarapax et Ramskoeldia ont parfois été remises en question. La monophylie de la famille spécifique des Hurdiidae est bien soutenue par plusieurs caractères dérivés (par exemple, région articulée distale de l'appendice frontal avec 5 podomères proximaux portant des endites subégaux), les Tamisiocarididae étant souvent suggérés comme son groupe frère.
-
Radiodonta
- ? Cucumericrus
- Caryosyntrips
-
Anomalocarida
- Paranomalocaris (placé dans Anomalocarididae par certaines études.)
- Laminacaris (placé dans Amplectobeluidae par certaines études.)
-
Anomalocarididae
- Anomalocaris (certaines espèces peuvent être placées dans d'autres familles.)
-
Amplectobeluidae
- Lyrarapax (position mise en doute par certaines études.)
- Amplectobelua
- Ramskoeldia (position mise en doute par certaines études.)
- Tamisiocarididae
- Hurdiidae
La première analyse phylogénétique approfondie de Radiodonta a été menée par Vinther et al . en 2014, et il a été élargi par Cong et al . plus tard cette année-là, par l'ajout de Lyrarapax unguispinus . L'analyse a été encore modifiée en 2015 par Van Roy et al. avec des caractères modifiés et l'inclusion de Cucumericrus decoratus et Aegirocassis benmoulai .
|


| Espèce | Descripteurs | Année nommée | Famille | Âge | Emplacement | Appendice frontal |
|---|---|---|---|---|---|---|
| Cucumericrus decoratus | Hou, Bergström et Ahlberg | 1995 | Inconnue | |||
| Caryosyntrips serratus | Daley et Budd | 2010 | Wuliuan - drumien |
|

|
|
| Caryosyntrips camurus | Pates et Daley | 2017 | Wuliuan |
|

|
|
| Caryosyntrips durus | Pates et Daley | 2017 | Tambour |
|

|
|
| Paranomalocaris multisegmentalis | Wang, Huang et Hu | 2013 |
|

|
||
| Chimère Laminacaris | Guo, Pates, Cong, Daley, Edgecombe, Chen et Hou | 2018 |
|

|
||
| Anomalocaris canadensis | Feuilles blanches | 1892 | Anomalocarididae |

|
||
| Anomalocaris pennsylvanica | Resser | 1929 |

|
|||
| Anomalocaris kunmingensis | Wang, Huang et Hu | 2013 | ||||
| Anomalocaris magnabasis | Pates, Daley, Edgecombe, Cong et Lieberman | 2019 |

|
|||
| Anomalocaris saron | Hou, Bergström et Ahlberg | 1995 |

|
|||
| Anomalocaris briggsi | Nedin | 1995 | Tamisiocarididae |
|
|
|
| Ramskoeldia platyacantha | Cong, Edgecombe, Daley, Guo, Pates et Hou | 2018 | Amplectobeluidae |

|
||
| Ramskoeldia consimilis | Cong, Edgecombe, Daley, Guo, Pates et Hou | 2018 | Amplectobeluidae |

|
||
| Lyrarapax unguispinus | Cong, Ma, Hou, Edgecombe et Strausfield | 2014 | Amplectobeluidae | Cambrien Étape 3 |
|

|
| Lyrarapax trilobus | Cong, Daley, Edgecombe, Hou et Chen | 2016 | Amplectobeluidae | Cambrien Étape 3 |
|

|
| Amplectobelua symbrachiata | Hou, Bergström et Ahlberg | 1995 | Amplectobeluidae |

|
||
| Amplectobelua stephenensis | Daley et Budd | 2010 | Amplectobeluidae |

|
||
| Tamisiocaris borealis | Daley et Peel | 2010 | Tamisiocarididae |
|

|
|
| Ursulinacaris grallae | Pates, Daley et Butterfield | 2019 | Hurdiidae | Wuliuan |
|

|
| Schinderhannes Bartelsi | Kühl, Briggs et Rust | 2009 | Hurdiidae | Emsian |
|
|
| Stanleycaris Hirpex | Pates, Daley et Ortega-Hernández | 2018 | Hurdiidae |
|
||
| Peytoia nathorsti | Walcott | 1911 | Hurdiidae |

|
||
| Peytoia infercambriensis | Lendzion | 1975 | Hurdiidae | Cambrien Étape 3 |
|
|
| Aegirocassis benmoulai | Van Roy, Daley et Briggs | 2015 | Hurdiidae | Trémadocien |
|

|
| Hurdia victoria | Walcott | 1912 | Hurdiidae | Wuliuan - drumien |
|

|
| Hurdia triangulata | Walcott | 1912 | Hurdiidae | Wuliuan |
|
|
| Cambroraster falcatus | Moysiuk et Caron | 2019 | Hurdiidae | Wuliuan |
|

|
| Pahvantia hastata | Robison et Richards | 1981 | Hurdiidae | Tambour |
|
|
| Cordaticaris striatus | Sun, Zeng et Zhao | 2020 | Hurdiidae | Tambour |
|
|
| Zhenghecaris shankouensis | Vanner, Chen, Huang, Charbonnier et Wang | 2006 | Hurdiidae |
|
Inconnue |
Histoire
L'histoire des radiodontes est complexe. Les spécimens incomplets appartenant à différentes parties du corps de la même espèce avaient été historiquement interprétés comme appartenant à différentes espèces et même à différents phylums. Avant leur reconnaissance en tant que groupe, les spécimens de radiodontes avaient été attribués à cinq phylums différents: Porifera , Cnidaria , Echinodermata , Annelida et Arthropoda .
Les premiers spécimens de radiodontes connus ont été recueillis dans les gisements de trilobites du mont Stephen par Richard G. McConnell de la Commission géologique du Canada en 1886 ou 1888. Ces spécimens ont été nommés Anomalocaris canadensis en 1892 par le paléontologue de GSC Joseph Whiteaves . Whiteaves a interprété les spécimens, maintenant connus pour être des appendices frontaux isolés, comme l'abdomen d'un crustacé phyllocaridé . Des spécimens de radiodonte supplémentaires ont été décrits en 1911 par Charles Walcott . Il a interprété un cône oral isolé, qu'il a nommé Peytoia , comme une méduse, et un spécimen mal conservé mais relativement complet, qu'il a appelé Laggania , comme un holothurie. En 1912, Walcott nomma Hurdia sur la base d'un élément h isolé, qu'il interpréta comme la carapace d'un crustacé. Un élément p de Hurdia a été nommé Proboscicaris en 1962 et interprété comme la carapace d'un arthropode bivalve.
La Commission géologique du Canada a entrepris une révision des fossiles de schiste de Burgess en 1966, sous la supervision du paléontologue de l' Université de Cambridge, Harry B. Whittington . Cette révision conduirait à terme à la découverte du plan complet du corps du radiodontisme. En 1978, Simon Conway Morris a reconnu que les pièces buccales de Laggania étaient Peytoia -comme, mais il a interprété cela comme une preuve qu'il était un fossile composite constitué d'une Peytoia méduses et une éponge. En 1979, Derek Briggs a reconnu que les fossiles d' Anomalocaris étaient des appendices, pas des abdomens, mais les a interprétés comme des jambes qui marchent. Ce n'est qu'en 1985 que la vraie nature des fossiles d' Anomalocaris , Laggania et Peytoia a été reconnue, et ils ont tous été attribués à un seul genre, Anomalocaris . Par la suite, il a été reconnu qu'Anomalocaris était une forme distincte des deux autres, ce qui a entraîné une scission en deux genres, dont le dernier a été nommé différemment Laggania et Peytoia jusqu'à ce qu'il soit déterminé que Peytoia avait la priorité. Il a été reconnu plus tard que certains des fossiles attribués à ces taxons appartenaient à une autre forme, qui a été reconnue comme portant une carapace composée d' éléments Hurdia et Proboscicaris . Enfin, en 2009, ces spécimens ont été redécrits comme Hurdia . Radiodonta était à l'origine considéré comme contenant une seule famille, Anomalocarididae, mais il a été divisé en quatre familles en 2014.